RÉSUMÉ
TLT-1 (Triggering Receptor Expressed on Myeloid cells-like Transcript-1, TREML1) est un récepteur de la super-famille des immunoglobulines exclusivement exprimé par les mégacaryocytes et les plaquettes. Localisé dans les granules α des plaquettes au repos, il est rapidement externalisé à la surface membranaire lors de leur activation. Initialement considéré comme un simple marqueur d’activation, TLT-1 est aujourd’hui reconnu comme un régulateur majeur de la thrombo-inflammation à l’interface entre hémostase, thrombose et immunité innée. Les études expérimentales montrent que TLT-1 favorise la stabilisation du thrombus en renforçant les interactions dépendantes du fibrinogène et en modulant les voies de signalisation intracellulaire impliquées dans l’agrégation plaquettaire. Son expression rapide et sa distribution étendue au sein du thrombus en font un marqueur plus sensible que la P-sélectine pour détecter les états précoces ou intermédiaires d’activation plaquettaire. TLT-1 participe également au maintien de l’intégrité vasculaire au cours des réponses inflammatoires systémiques. Sa forme soluble (sTLT-1), libérée après activation plaquettaire, constitue un biomarqueur circulant robuste de l’activation plaquettaire et de l’immunothrombose. Des concentrations élevées de sTLT-1 ont été rapportées dans plusieurs pathologies thrombo-inflammatoires sévères, notamment le sepsis, la coagulation intravasculaire disséminée, la prééclampsie et certaines maladies cardiovasculaires inflammatoires. Ainsi, TLT-1 apparaît aujourd’hui comme une cible thérapeutique prometteuse pour moduler la thrombo-inflammation tout en préservant l’hémostase physiologique.MOTS CLÉS
activation plaquettaire, hémostase, thrombo-inflammation, TLT-1
ABSTRACT
Triggering Receptor Expressed on Myeloid Cells-Like Transcript 1 (TLT-1, TREML1) is an immunoglobulin superfamily receptor exclusively expressed in megakaryocytes and platelets. Stored in platelet α-granules under resting conditions, it is rapidly translocated to the plasma membrane upon platelet activation. Initially considered a simple platelet activation marker, TLT-1 is now recognized as a major regulator of thrombo-inflammation at the crossroads between hemostasis, thrombosis, and innate immunity. Experimental studies demonstrate that TLT-1 promotes thrombus stabilization by strengthening fibrinogen-dependent interactions and modulating intracellular signaling pathways involved in platelet aggregation. Its rapid surface exposure and broad distribution within the thrombus make TLT-1 a more sensitive marker than P-selectin for detecting early or intermediate stages of platelet activation. TLT-1 also contributes to vascular integrity during systemic inflammatory responses. Its soluble form (sTLT-1), released following platelet activation, has emerged as a robust circulating biomarker of platelet activation and immunothrombosis. Elevated plasma levels of sTLT-1 have been reported in several thrombo-inflammatory disorders, including sepsis, disseminated intra-vascular coagulation, preeclampsia, and inflammatory cardiovascular diseases. Collectively, these findings identify TLT-1 as a promising therapeutic target for the modulation of thrombo-inflammation while preserving physiological hemostasis.
KEYWORDS
hemostasis, platelet activation, thrombo-inflammation, TLT-1
INTRODUCTION
Longtemps considérées comme de simples effecteurs de l’hémostase, les plaquettes sont aujourd’hui reconnues comme de véritables cellules immunitaires circulantes capables de participer activement aux processus inflammatoires et à l’immunité innée (1). Outre leur rôle central dans la formation du thrombus, elles interagissent avec les leucocytes, les cellules endothéliales et les composants du complément afin de coordonner les réponses vasculaires et inflammatoires. Les granules α plaquettaires contiennent ainsi de nombreuses protéines impliquées non seulement dans l’hémostase, mais également dans l’inflammation, notamment des cytokines ou chimiokines telles que le TGF-β ou le PF4, et des molécules d’adhésion. Dans ce contexte, le concept de thrombo-inflammation, ou immunothrombose, a émergé pour décrire les interactions étroites entre coagulation et immunité innée. Plusieurs récepteurs immunitaires exprimés par les plaquettes participent à cette interface fonctionnelle. Parmi eux, TLT-1 occupe une place particulière en raison de sa spécificité d’expression dans la lignée mégacaryocytaire et de sa cinétique d’activation extrêmement rapide. TLT-1 appartient à la famille des récepteurs TREM (Triggering Receptor Expressed on Myeloid Cells) (2). Contrairement à TREM-1, exprimé principalement par les cellules myéloïdes, TLT-1 est restreint aux mégacaryocytes et aux plaquettes (3,4). Il constitue l’un des récepteurs contenant des motifs ITIM (Immunoreceptor Tyrosine-based Inhibitory Motif) les plus abondants dans les plaquettes humaines et murines (5-9). Initialement décrit comme un marqueur d’activation plaquettaire, il est désormais considéré comme un régulateur fonctionnel majeur de la thrombo-inflammation. Les données expérimentales montrent qu’il participe à la stabilisation du thrombus, aux interactions plaquettes-leucocytes et au maintien de l’intégrité vasculaire au cours des processus inflammatoires. Par ailleurs, sa forme soluble (sTLT-1), libérée après activation plaquettaire, apparaît comme un biomarqueur prometteur des états thrombo- inflammatoires sévères. Cette revue synthétise les connaissances actuelles concernant les implications physiopathologiques de TLT-1 dans l’activation plaquettaire et la thrombo-inflammation.
STRUCTURE
TLT-1 est codé par le gène TREML1, localisé sur le chromosome 6 chez l’homme et le chromosome 13 chez la souris. Il appartient à la superfamille des immuno-globulines et possède un domaine extracellulaire de type immunoglobuline variable, une région transmembranaire et une courte queue cytoplasmique contenant des motifs de signalisation intracellulaire ITIM et ITSM (Immunoreceptor Tyrosine-based Switch Motif). Trois isoformes issues de variants d’épissage du TLT-1 ont été identifiées qui ne diffèrent que par leurs queues intracellulaires. La forme complète comporte une longue queue intracellulaire de 127 acides aminés contenant deux résidus tyrosine conservés (Y245 et Y281) intégrés dans un motif ITIM classique C-terminal et un motif ITSM non-classique proximal à la membrane (TxYxxL et VxYxxV) (10). Ces domaines permettent le recrutement des protéines tyrosine-phosphatases SHP-1 et SHP-2 (Src Homology containing tyrosine phosphatases) et la propagation d’un signal intracellulaire inhibiteur. Une région riche en prolines (PxxP motifs) permet quant à elle une interaction avec des protéines porteuses de domaines SH3 (11,12). Une seconde isoforme, variante d’épissage plus courte (TLT-1sp), possède une queue intracellulaire de 18 acides aminés et dépourvue de motifs ITIM/ITSM. En lieu et place, cette isoforme contient un motif de tri des récepteurs dileucine similaire à celui des molécules MHC de classe II à chaîne variable (10). Une troisième isoforme correspond à une forme soluble (sTLT-1) constituée uniquement du domaine extracellulaire. L’existence de ces variants suggère des fonctions distinctes, indépendantes ou non de la signalisation intracellulaire (2,12). Contrairement aux membres classiques de la famille TREM exprimés principalement par les cellules myéloïdes, TLT-1 ne s’associe pas au co-récepteur DAP12 et son expression est strictement limitée à la lignée mégacaryocytaire et plaquettaire (Figure 1). Dans les plaquettes au repos, TLT-1 est majoritairement localisé dans les granules α. Après activation par différents agonistes tels que la thrombine, le collagène, l’ADP ou les stimuli inflammatoires, il est rapidement transloqué vers la membrane plasmique (Figure 1). Cette externalisation précoce fait de TLT-1 un marqueur particulièrement sensible de l’activation plaquettaire aiguë, détectable en surface plus précocement que la P-sélectine (Figure 2). La forme soluble de TLT-1 peut également être libérée dans la circulation après activation plaquettaire, par clivage protéolytique de la forme membranaire par des métalloprotéinases (10,11). Parmi les différentes substances libérées sous forme soluble par les plaquettes activées, sTLT-1 est en proportion équivalente à certaines glycoprotéines plaquettaires essentielles telles que GPIb, GPIIbIIIa et GPV, suggérant un rôle fonctionnel important (12).
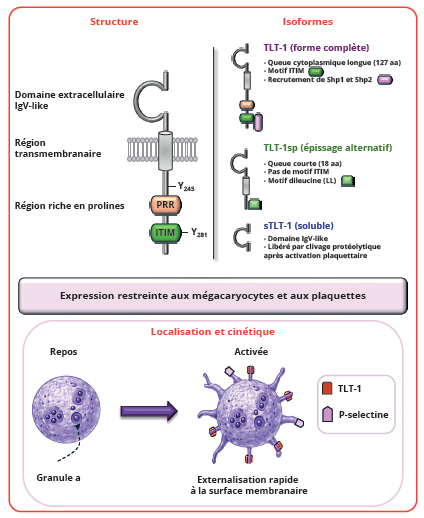
Figure 1 : Organisation structurale de TLT-1, principales isoformes et localisation cellulaire.
Figure 1: Structural organization of TLT-1, major isoforms, and cellular localization.
TLT-1 est spécifiquement exprimé dans les granules α des mégacaryocytes et les plaquettes, puis est rapidement exposé à la membrane plasmique après activation plaquettaire.
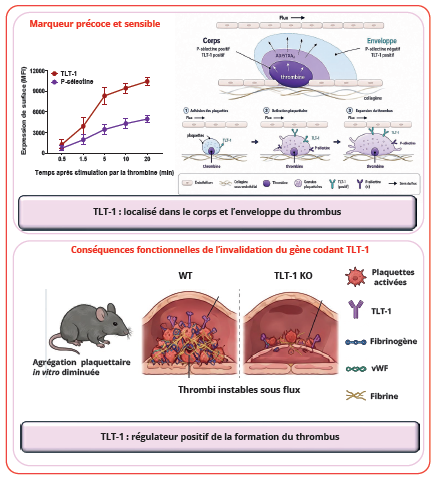
Figure 2 : Implication de TLT-1 dans l’activation plaquettaire et la thrombose.
Figure 2: The role of TLT-1 in platelet activation and thrombosis.
TLT-1 favorise l’agrégation plaquettaire et contribue à la croissance ainsi qu’à la stabilisation du thrombus.
TLT-1 ET ACTIVATION PLAQUETTAIRE
Un marqueur précoce de l’activation plaquettaire
TLT-1 est colocalisé avec la P-sélectine dans les granules α des plaquettes au repos. Cependant, après activation, les deux protéines présentent des distributions distinctes (11,13). Alors que la P-sélectine est principalement externalisée à la surface membranaire afin d’assurer les interactions adhésives avec les leucocytes et l’endothélium (14), TLT-1 présente une distribution plus diffuse incluant une région centrale proche de la bande marginale plaquettaire. La P-sélectine est une protéine « cargo » dont la fonction d’adhésion est portée par sa portion extracellulaire.
Ainsi, elle ne comporte qu’une courte queue cytoplasmique de 35 résidus dont la délétion n’altère pas sa trans- location membranaire lors de l’activation plaquettaire (14). Par conséquent, il est probable que la différence entre les domaines cytoplasmiques des deux isoformes de TLT-1 soit responsable de l’adressage différentiel de TLT-1 au cours de l’activation plaquettaire, et que la fonction associée aux deux isoformes en diffère. Plusieurs travaux ont montré que TLT-1 constitue un marqueur plus sensible de l’activation plaquettaire que la P-sélectine, longtemps considérée comme le gold standard. L’expression membranaire de TLT-1 apparaît plus rapidement et avec une intensité supérieure à celle de la P-sélectine après stimulation plaquettaire in vitro (Figure 2). Dans les modèles expérimentaux de thrombose in vivo, TLT-1 est détecté non seulement dans le cœur hautement activé du thrombus mais également dans la région périphérique constituée de plaquettes partiellement activées. À l’inverse, la P-sélectine reste confinée au cœur hautement activé du thrombus (Figure 2). Cette distribution particulière suggère que TLT-1 permettrait de détecter des états précoces à intermédiaires d’activation plaquettaire difficilement détectables par les marqueurs conventionnels (13,15,16). Ainsi, TLT-1 pourrait représenter un marqueur intéressant pour le dépistage des états prothrombotiques précoces.
Interaction avec le fibrinogène et agrégation plaquettaire
TLT-1 joue également un rôle fonctionnel dans l’agrégation plaquettaire. L’analyse structurale du domaine extra- cellulaire du TLT-1 a permis de mettre en évidence plusieurs sites de liaison, notamment pour le fibrinogène et le facteur Willebrand (17). Ceci a permis de positionner TLT-1 comme un régulateur de l’hémostase facilitant la formation d’agrégats plaquettaires. Sa forme soluble favorise l’agrégation induite par de faibles concentrations d’agonistes en stabilisant les interactions plaquettes- plaquettes dépendantes du fibrinogène. Cette interaction semble indépendante du motif RGD classiquement impliqué dans la liaison du fibrinogène à l’intégrine αIIbβ3. L’ajout de sTLT-1 recombinant augmente l’agrégation induite par des concentrations suboptimales de collagène, d’ADP et d’un analogue du TXA2 (U46619) (12). À l’inverse, le blocage de TLT-1 par des anticorps dirigés contre le domaine extracellulaire inhibe l’agrégation induite par de faibles quantités de thrombine, probablement en bloquant la liaison de son ligand sur TLT-1 (18), suggérant que TLT-1 agit comme un amplificateur secondaire de l’activation. TLT-1 interagit également avec les protéines de la famille ezrin/radixin/moesin (ERM) (12), établissant potentiellement un lien entre le fibrinogène extracellulaire et le cytosquelette plaquettaire. Cette propriété pourrait contribuer à la stabilisation mécanique du thrombus et à l’organisation de réarrangements cytosquelettiques nécessaires à l’adhésion et à l’étalement plaquettaires. Des études plus récentes ont montré que TLT-1 interagit avec une forte affinité avec le facteur Willebrand, proposé par conséquent comme ligand physiologique de TLT-1 (17). Le blocage des interactions du TLT-1 avec ses ligands à l’aide d’anticorps bloquants réduit la formation d’agrégats plaquettaires sur une matrice de collagène de type 1 soumise à des forces de cisaillement élevées, soulignant l’importance potentielle de TLT-1 dans les thromboses artérielles. Le rôle fonctionnel de TLT-1 a été confirmé dans des modèles murins déficients pour TREML1 (Figure 2). Les souris knock-out (KO) présentent ainsi un déficit de l’agrégation dépendante de l’ADP et du thromboxane A2 (12). Mais, le blocage de TLT-1 n’altère ni le changement de forme ni la dégranulation des plaquettes. Ainsi TLT-1 interviendrait plutôt au cours de l’hémostase primaire dans les phénomènes d’auto-amplification de l’agrégation après relargage du contenu granulaire. TLT-1, tant dans sa forme membranaire que dans sa forme soluble, serait capable de lier le fibrinogène libéré par les plaquettes activées, potentiellement de concert avec GPIIbIIIa, afin de faciliter la liaison du fibrinogène sur les plaquettes et/ou augmenter la stabilité de l’agrégat, en créant par exemple des ponts intermoléculaires (10). L’activation de GPIIbIIIa par le fibrinogène entraîne le recrutement et l’activation de tyrosine-kinases de la famille Src et Syk (19). C’est la première phase d’un ensemble de signaux outside-in conduisant in fine à des réarrangements du cytosquelette et à la polymérisation des filaments d’actine, processus important pour l’étalement plaquettaire sur les matrices extracellulaires, l’adhésion plaquettaire, la stabilité de l’agrégat plaquettaire et pour certains phénomènes plus tardifs comme la rétraction du caillot après la formation du thrombus (20). Finalement, l’absence de TLT-1 chez les souris KO se traduit par une instabilité des thrombi formés sous flux, ce résultat soutenant le rôle clé de TLT-1 pour le maintien de leur intégrité. Ainsi, TLT-1 apparaît comme un régulateur positif de la stabilité thrombotique plutôt qu’un initiateur primaire de l’activation plaquettaire.
TLT-1 ET THROMBO-INFLAMMATION
Interface entre hémostase et immunité innée
Les plaquettes jouent un rôle majeur dans l’immunité innée en détectant les signaux inflammatoires et en modulant les interactions leucocytaires et endothéliales. Dans ce contexte, TLT-1 apparaît comme un acteur important de l’immunothrombose. Les concentrations plasmatiques de sTLT-1 augmentent significativement au cours du sepsis et sont corrélées avec la présence d’une coagulation intravasculaire disséminée (CIVD) (21-23). Ces travaux ont également souligné l’intérêt potentiel du sTLT-1 comme biomarqueur diagnostique et pronostique dans les pathologies inflammatoires sévères. Les concentrations circulantes de sTLT-1 sont également corrélées à la gravité clinique, à l’activation systémique des plaquettes et au risque de dysfonction multiviscérale. Chez les souris TLT-1 KO exposées au lipopolysaccharide (LPS), une augmentation des concentrations de TNF-α, des D-dimères et de la mortalité est observée, avec des hémorragies inflammatoires sévères (24). Ces données suggèrent que TLT-1 participe au maintien de l’intégrité vasculaire au cours de l’inflammation systémique en stabilisant les interactions plaquettaires au niveau de la microcirculation.
Régulation de la thrombo-inflammation
Plusieurs données suggèrent également que TLT-1 intervient dans les interactions entre plaquettes et leucocytes, notamment avec les neutrophiles, acteurs centraux de l’immunité innée et de l’immunothrombose (Figure 3a). Lors des réponses inflammatoires systémiques, les plaquettes activées adhèrent aux neutrophiles via différentes molécules d’adhésion et favorisent leur activation fonctionnelle. Dans ce contexte, l’externalisation rapide de TLT-1 à la surface des plaquettes pourrait contribuer à stabiliser ces interactions cellulaires au sein de la microcirculation inflammatoire. L’un des mécanismes majeurs de l’immunothrombose repose sur la formation des NETs (Neutrophil Extracellular Traps) (25), structures extra-cellulaires composées d’ADN décondensé, d’histones et de protéines granulaires antimicrobiennes libérées par les neutrophiles activés. Bien que les NETs participent initialement à la défense anti-infectieuse en piégeant les pathogènes, leur production excessive favorise également les lésions endothéliales, l’activation de la coagulation et la propagation du thrombus.
L’implication potentielle de TLT-1 dans la formation des NETs renforce l’hypothèse de son rôle central dans les mécanismes d’immunothrombose, bien qu’ils demeurent encore incomplètement caractérisés. Dans ce contexte, plusieurs observations suggèrent que TLT-1 pourrait agir comme un modulateur important de la communication plaquettes–neutrophiles. Les concentrations élevées de sTLT-1 mesurées au cours du sepsis et des états inflammatoires sévères coïncident avec une activation accrue des neutrophiles et une augmentation de la formation des NETs (26). Sur le plan mécanistique, plusieurs hypothèses sont actuellement envisagées. TLT-1 pourrait favoriser le recrutement localisé des neutrophiles au sein du thrombus, moduler leur activation via des interactions directes ou indirectes avec le fibrinogène et le facteur Willebrand, ou encore influencer la stabilisation des NETs dans le réseau fibrinoplaquettaire. TLT-1 pourrait ainsi agir comme un régulateur de l’équilibre entre hémostase protectrice et inflammation excessive, limitant les lésions vasculaires tout en maintenant une réponse thrombotique locale efficace.
TLT-1 : BIOMARQUEUR DES ÉTATS PROTHROMBOTIQUES
L’ensemble des données expérimentales indique que TLT-1 favorise la formation du thrombus en augmentant l’efficacité des interactions dépendantes du fibrinogène. Contrairement aux récepteurs d’adhésion classiques comme l’intégrine αIIbβ3, TLT-1 semble agir comme un amplificateur secondaire de l’activation plaquettaire. Sa distribution diffuse dans le thrombus, y compris dans les zones périphériques faiblement activées, suggère une contribution importante à la stabilité thrombotique.
Plusieurs études cliniques ont rapporté une augmentation des concentrations plasmatiques de sTLT-1 dans différents contextes pathologiques, notamment le sepsis et la CIVD, la prééclampsie et certaines maladies cardiovasculaires thrombogènes (Figure 3b). Dans la prééclampsie, l’expression de TLT-1 et les concentrations circulantes de sTLT-1 sont augmentées parallèlement aux marqueurs classiques d’activation plaquettaire (27). Certaines études suggèrent même que TLT-1 pourrait être un marqueur plus discriminant que la P-sélectine dans ce contexte. Dans le sepsis, l’élévation du sTLT-1 semble corrélée à l’intensité de l’inflammation et à l’activation systémique des plaquettes. Cette augmentation pourrait refléter la participation des plaquettes aux mécanismes d’immunothrombose. Les études expérimentales suggèrent également que TLT-1 contribue à la stabilité du thrombus dans les thromboses artérielles. Les modèles murins déficients en TLT-1 présentent une protection partielle contre les occlusions thrombotiques excessives. Ces observations ouvrent donc des perspectives thérapeutiques importantes puisque le ciblage de TLT-1 pourrait permettre de limiter certaines réponses thrombotiques pathologiques tout en préservant une hémostase physiologique suffisante.
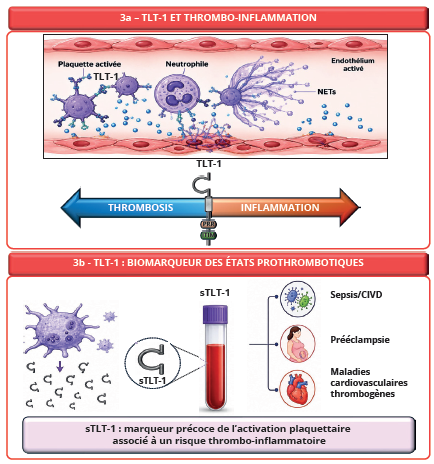
Figure 3 : Rôle de TLT-1 dans la thrombo-inflammation et potentiel biomarqueur d’états prothrombotiques.
Figure 3: The role of TLT-1 in thromboinflammation and its potential as a biomarker of prothrombotic states.
La forme soluble sTLT-1 constitue un marqueur de l’activation plaquettaire associé à plusieurs états prothrombotiques et thrombo-inflammatoires.
PERSPECTIVES THÉRAPEUTIQUES
La spécificité plaquettaire de TLT-1 en fait une cible thérapeutique particulièrement intéressante. Contrairement aux antiagrégants plaquettaires actuellement utilisés, dont l’efficacité s’accompagne souvent d’un risque hémorragique significatif, le ciblage de TLT-1 pourrait permettre de moduler sélectivement les mécanismes thrombo-inflammatoires pathologiques tout en préservant l’hémostase physiologique. Dans cette optique, une avancée récente majeure a conduit au développement d’un anticorps bispécifique innovant destiné au traitement prophylactique des maladies hémorragiques héréditaires, en particulier la thrombasthénie de Glanzmann (Figure 4) (28). En effet, cet anticorps est capable de se lier avec une forte affinité au FVIIa endogène via l’un de ses bras, et au récepteur TLT-1 exprimé à la surface des plaquettes activées via son second bras. Cette stratégie présente un intérêt mécanistique important. Grâce à la longue demi-vie de l’anticorps, des complexes circulants HMB-001/FVIIa s’accumulent dans le plasma, permettant de prolonger considérablement la disponibilité systémique du FVIIa endogène et favorisant une hémostase localisée au niveau des plaquettes activées exprimant TLT-1. Cette étude illustre ainsi le potentiel translationnel du ciblage de TLT-1 pour développer des approches thérapeutiques capables de restaurer une hémostase efficace tout en limitant le risque de thrombose systémique excessive (28).
Plusieurs questions demeurent néanmoins ouvertes, notamment concernant l’identification complète des partenaires moléculaires de TLT-1, les voies de signalisation intracellulaire activées, son rôle exact dans la thrombo-inflammation et les conséquences d’un ciblage pharmacologique chronique. Une meilleure compréhension de ces mécanismes sera indispensable avant toute application clinique ou thérapeutique.
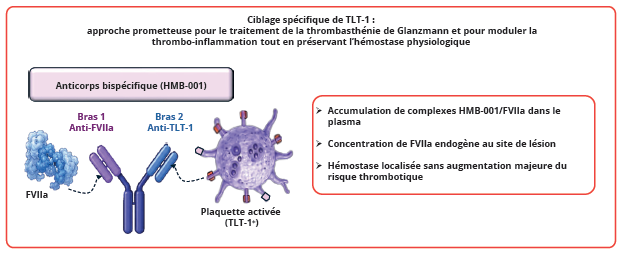
Figure 4 : Exploitation thérapeutique de TLT-1 comme cible plaquettaire spécifique.
Figure 4: Therapeutic use of TLT-1 as a specific platelet target.
Le ciblage de TLT-1 permet d’améliorer l’hémostase ou de moduler la thrombo-inflammation tout en limitant le risque thrombotique systémique.
CONCLUSION
TLT-1 est aujourd’hui reconnu comme un récepteur plaquettaire majeur impliqué dans l’activation plaquettaire, la stabilisation du thrombus et les interactions entre coagulation et inflammation. Son expression rapide à la surface des plaquettes activées, sa capacité à lier le fibrinogène et son implication dans le maintien de l’intégrité vasculaire en font un acteur clé de la thrombo-inflammation. La forme soluble de TLT-1 apparaît également comme un biomarqueur prometteur des états prothrombotiques et inflammatoires systémiques. Bien que plusieurs aspects de sa biologie restent encore à élucider, les données actuelles positionnent TLT-1 comme une cible thérapeutique potentielle dans les pathologies cardiovasculaires et inflammatoires associées à l’immunothrombose.
POINTS CLÉS À RETENIR
• TLT-1 est un récepteur spécifiquement exprimé par les mégacaryocytes et les plaquettes, rapidement externalisé lors de l’activation plaquettaire.
• TLT-1 constitue un marqueur plus précoce et sensible de l’activation plaquettaire que la P-sélectine.
• En se liant au fibrinogène, TLT-1 favorise la stabilisation du thrombus et participe au maintien de l’intégrité vasculaire.
• TLT-1 joue un rôle central dans la thrombo-inflammation en reliant activation plaquettaire, thrombose et immunité innée.
• La forme soluble sTLT-1 représente un biomarqueur prometteur des états thrombo-inflammatoires sévères et une cible thérapeutique potentielle.
Liens d’intérêts : l’auteur déclare ne pas avoir de lien d’intérêt en rapport avec cet article.